
Brief communication
The role of commotion in the egg as means of feto-maternal communication.
A suitable temperature is crucial for a successful embryonic development. The needed temperature is most often different from the ambient temperature and bird’s thermoregulatory abilities fully mature only after hatching. Therefore all avian embryos (except megapodes) are dependent on the assistance of an incubating parent (Webb 1987) . During natural incubation there are great costs associated with incubation for the incubating parents. He or she must find a working balance between their own need for energy with the thermal need of the embryos (Conway & Martin 2000) . Consequently a mean for the fetus to communicate its well-being would be beneficial for both it self and the incubating parent. Some changes in embryonic activity indicate a need for heating assistance and can therefore be possible means of embryo-parent communication (Abraham & Evans 1999) . The most commonly studied mean of feto-maternal interaction is communication through vocalization. The results form these studies have been contradictory (Bugden & Evans 1999) .
The aim of this study was to test, not only vocalization but also some other of the possible means of communication and if detected responses could be linked to any particular temperatures. The variables of interest are associated with embryonic vocalizations, embryonic movements, ventilation frequency and oxygen consumption. The studies were conducted on eggs from broiler chickens (Gallus gallus domesticus).
Eggs of three different developmental stages were used in the study: 18 days old (18), internally pipped (IP) and externally pipped (EP). Each egg was used once and exposed to five different temperature intervals T1-T5 ranging from 40◦C down to 30◦C. All variables were recorded for 15min in each of the five temperature intervals.
Both differences between the ages and the ration of maximum peak trends within the ages were tested for all the different variables. The maximum trend response is characterized by an increase followed by a decrease (see example on ventilation frequency in figure 1). The maximum peak trend will occur in a 1:4 ratio if no temperature related responses occurred.
All three ages showed linear responses in oxygen consumption to lowered temperatures. The slopes of the response line were significantly steeper in embryos of age EP and IP than it was in embryos age 18. No indications of an increased metabolic rate as a response to decreased temperatures where seen. No significant maximum peak trend (see figure 1) was seen in any of the tested vocalization variables. Phrase duration was significantly longer in EP than in IP embryos. No other differences between the ages where found.
A significant higher number of embryos showed a peak described as a maximum peak trend on several of the tested movement variables and on ventilation frequency. The peak occurred during the third temperature interval (T3) when the temperature was between 35.0ºC and 35.5ºC. The results suggests that the temperature threshold important as an embryonic activity trigger is the lower limit of optimal development which is shown to occurs, for broiler chickens, just below 36◦C (Conway & Martin 2000) .
Oxygen consumption is a reliable indicator of the thermal status but changes are linear and not linked to any particular temperatures and the possibility that the incubating parent can detect theses very feeble changes seems extremely unlikely.
Embryonic vocalizations are not suitable as indicators of decreasing temperatures for broiler chickens (Gallus gallus domesticus) embryos and are therefore most certainly not of importance as a mean to communicate the thermal status of the embryo to the incubating parent.
Increased embryonic movement activity and ventilation frequencies are in the study shown to indicate that ambient temperature has gone below the threshold temperature of optimal development and consequently can be used as means of fetal-maternal communication in broiler chickens (Gallus gallus domesticus).
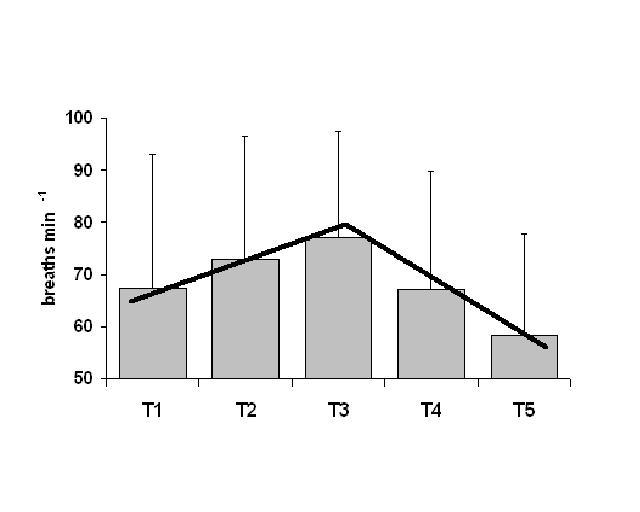
Figure 1. Breathing frequency of internally pipped embryos
(IP) during five different temperature intervals (T1-T5). Columns are
means of 10 sets of embryos and bars are standard deviation. The two
lines (T1 – T3 with a positive slope and T3 – T5 with a negative slope)
show the occurrence of a maximum peak trend.
References:
Abraham, C. L. & Evans, R. M. (1999). Metabolic costs of heat solicitation calls in relation to thermal need in embryos of American white pelicans. Animal Behaviour 57, 967-975.
Bugden, S. C. & Evans, R. M. (1999). The development of a vocal thermoregulatory response to temperature in embryos of the domestic chicken. Wilson Bulletin 111, 188-194.
Conway, C. J. & Martin, T. E. (2000). Effects of ambient temperature on avian incubation behavior. Behavioral Ecology 11, 178-188.
Webb, D. R. (1987). Thermal tolerance of avian embryos: a review. The Condor 89, 874-898.
Responsible for this page:
Director of undergraduate studies Biology
Last updated:
04/28/06