
Discussion
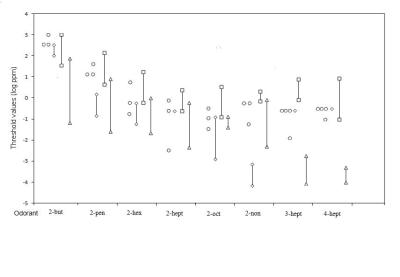
The results of the present study demonstrate a well-developed olfactory sensitivity of spider monkeys for aliphatic ketones. These findings are in line with earlier studies that used structurally related monomolecular substances such as aliphatic esters, alcohols and aldehydes, carboxylic acids, monoterpenes, steroids, thiazoles, thiols and indols, alkylpyrazines, aromatic aldehydes, green odours and proteinogenic amino acids. This lends further evidence to reject the old belief of primates, such as Spider monkeys, having a poor sense of smell. A significant negative correlation between olfactory sensitivity and carbon chain length was found in spider monkeys for the homologous 2-ketones. This finding corresponds with those found in squirrel monkeys and pigtail macaques. Further, when it comes to olfactory discrimination for aliphatic ketones it was found that among human test subjects it was significantly easier to discriminate longer carbon chains than shorter ones for both aliphatic ketones and acetic esters. A significant negative linear correlation between olfactory sensitivity and carbon chain length was also found in spider monkeys for carboxylic acids whereas the correlation found in pigtail macaques was U-shaped.This implies that the length of the carbon chain is a molecular feature that seems to be of importance for the sense of olfaction in primates in general and not only for aliphatic ketones. Furthermore, in agreement with the previous study on aliphatic ketones in squirrel monkeys and pigtail macaques no significant correlation between position of the functional group on the C7-ketones and the spider monkeys’ olfactory sensitivity was found. Thus, the position of the functional group may be of little importance for olfactory sensitivity in primates.
Three and, for a couple of odorants, four spider monkeys were used in the present study to determine the olfactory sensitivity for aliphatic ketones. This might appear as a small sample size. However, the individual threshold values of the spider monkeys were, for the majority of odorants, only differing by a dilution factor of three (for four of the odorants) or ten (for two of the odorants). Even though the olfactory sensitivity of the spider monkeys for two of the odorants differed by higher dilution factors between the most- and the least-sensitive animal, 33 respectively 300, the results appear trustworthy. Individual differences in olfactory sensitivity, sometimes even higher than those found in the present study, are regularly found in non-primate mammalian species i.e. in mice and occur not surprisingly in spider monkeys as well. The individual differences between the spider monkeys in this study are much lower than those found in human test subjects. This lends further support to the trustworthiness of the data.
The perception of odorants can change with age. Doty and Kamath reported (2014) humans to often experience a decrease in olfactory sensitivity when aging. However, Nordin et al. (2011) argue that even though a decrease of activity in olfactory brain structures is reported the olfactory sensitivity for different odorants is not automatically affected but can be as good as that found in young adults. The age of the spider monkeys in the present study ranged between 8 to 14 years old (8-, 11-, 14- and 14 years.). No obvious difference between the olfactory sensitivity due to the spider monkeys’ age was seen except that one of the 14 year old spider monkeys for the vast majority of odorants had the highest threshold values. On the contrary, the other 14 year old spider monkey demonstrated a very good olfactory sensitivity with aliphatic ketones in comparison with the rest of the spider monkeys. Since spider monkeys live for 20-27 years if healthy the animals in the present study are not considered too old to participate in olfactory sensitivity tests. Thus, even if individual differences in olfactory sensitivity can be influenced by age it may also be influenced by other factors i.e. in humans olfactory performance has been reported to be influenced by factors such as genetics, nutrition and well-being.
All of the spider monkeys were highly trained and three of them had participated in multiple studies using the same method prior to this study. Therefore, all of the test animals were well-acquainted with the tests and what they were supposed to do. This suggests that no test result was incorrect due to a spider monkey not knowing how to perform the task. Further, the number of trials (30) performed per dilution step also suggests that even if a spider monkey did make a mistake there were enough trials that combined would give a reliable result. If a spider monkey failed to reach criterion with a given dilution step by only one correct choice, then this dilution was tested again. This was done to double-check the trustworthiness of the results. All spider monkeys performed close to maximal and for the vast majority of odorants even made 100% correct choices when tested with the start dilution and thereafter, as the odorant got further diluted, a gradual decrease in their performance occurred until they reached the chance level. This leads me to believe that the only cue for the spider monkeys to decide for the box containing the food reward came from the paper strip with the odorant. Furthermore, the boxes were baited out of sight from the spider monkeys in a pseudorandom order thus there was no possibility for them to know which of the boxes was baited.
In the across-species comparison the data from humans are mean values. The data from the squirrel monkeys and the pigtail macaques are results from a previous study and the data from the spider monkeys are from the present study. For the human test subjects, the squirrel monkeys and the pigtail macaques the range between the highest and the lowest threshold values were used for the across-species comparison together with the threshold values from all spider monkeys in the present study. For the vast majority of odorants the threshold values for the four species were within the range of each other’s threshold values, that is: their ranges overlapped. For one of the odorants, 2-nonanone, the pigtail macaques even demonstrated lower threshold values than the three other species of primates. Furthermore, for two of the odorants, 3- and 4-heptanone, the human test subjects demonstrated lower threshold values than the other three species. This contradicts the conclusion drawn by Rouquier et al. in 2000 claiming hominoids to have the poorest sense of smell among primates followed by the Old World monkeys and then New World monkeys because of the increased fraction of OR pseudogenes during primate evolution. The results of the across-species comparison also contradict the hypothesis of a poorer sense of smell based on the relative size of olfactory brain structures which has decreased during primate evolution. Smith et al. (2007) argue that even though the anatomical features are beyond dispute it does not necessary correspond to a poorer sense of smell. The across-species comparison in this study suggests that anatomical and genetic features are of less importance for olfaction in primates.
Responsible for this page:
Director of undergraduate studies Biology
Last updated:
05/20/14