
Discussion
Sweetening Potency order
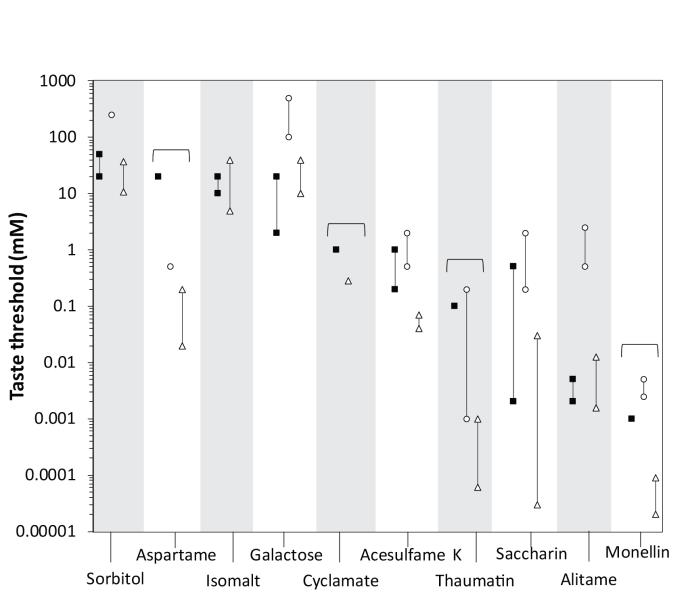
Figure 7 displays the spider monkeys’ taste preference thresholds, alongside with those of chimpanzees and humans. Even though the lowest taste preference threshold for spider monkeys was recorded for the sweet-tasting protein monellin, this value was recorded for only one of the four monkeys. Thus, the spider monkeys were generally the most sensitive to alitame, and the least sensitive to sorbitol. As alitame, sodium saccharine and acesulfame K were perceived by all spider monkeys and as the animals displayed taste preference thresholds at least one factor of 10 lower than that of sucrose, these sweet-tasting substances can be described as high-potency sweeteners in this species. As aspartame, isomalt and sorbitol have higher taste thresholds than sucrose, these substances can be described as low-potency sweeteners for the spider monkeys.
Similar to what was found for spider monkeys, alitame, sodium saccharin and acesulfame K also qualify as high-potency sweeteners in chimpanzees. Also similarly to the case of spider monkeys, sorbitol is also described as a low-potency sweetener for chimpanzees. Despite the similarities between the ranking order of sweetening potencies of spider monkeys and chimpanzees, no significant correlation was found when comparing the two.
Of the substances described as high-potency sweeteners for spider monkeys, only sodium saccharin holds the same description for humans. Similar to spider monkeys, sorbitol is described as a low-potency sweetener in humans. A comparison between the ranking order of sweetening potency of spider monkeys and humans showed a significant correlation between the order of ranking for the two species.
Between-species comparisons of taste preference thresholds
Artificial sweeteners
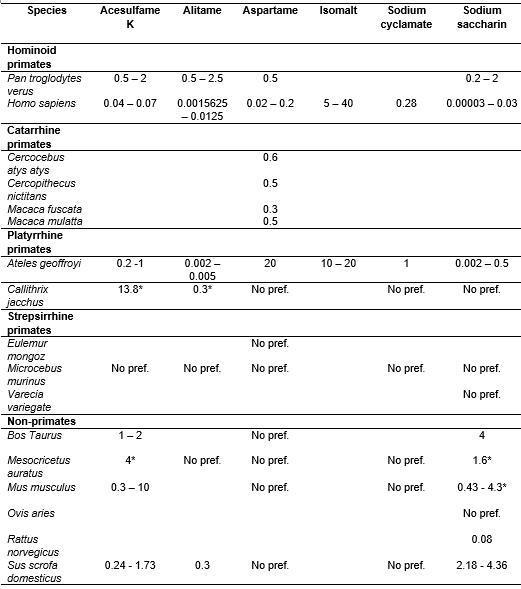
The spider monkeys detected 4 out of 6 artificial sweeteners. For the sweeteners which the spider monkeys clearly detect - such as acesulfame K, alitame, and sodium saccharine - their thresholds were generally lower than those of chimpanzees. This suggests a higher sensitivity to sweet-taste of spider monkeys compared to chimpanzees. There is not yet a reported taste preference threshold for isomalt for chimpanzees. The thresholds of the spider monkeys were either similar or higher than those of humans for the mentioned substances. However, it should be noted that the procedures used to determine taste preference thresholds in humans allow for the determination of taste detection thresholds rather than taste preference thresholds. Thus, the taste thresholds for humans represent the lowest concentration of a given substance that human subjects are able to detect but not necessarily prefer over water. As it is possible that animals detect lower concentrations of a given taste substance than the lowest one preferred over water (Spector 2003), it is then plausible to assume that spider monkeys may be able to detect acesulfame k concentrations as low as humans do.
Aspartame and sodium cyclamate are probably generally not perceived as sweet by the spider monkeys. This result for aspartame is in line with the notion that all Platyrrhine primates should be generally unable to detect its sweetness due to an underlying structural difference in the sweet-taste receptor of these species compared to Catarrhine primates. The individuals displaying a preference for both aspartame and sodium cyclamate over water most likely represent exceptional cases of allelic variation in the sweet-taste receptor's typical alleles.
Sweet-tasting proteins
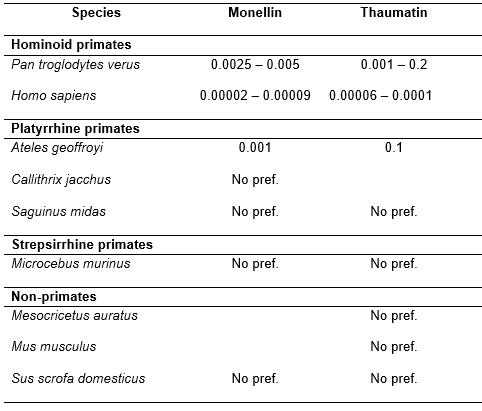
Spider monkeys are most likely generally unable to perceive the sweetness of monellin and thaumatin. These results are congruent with an explanatory theory, similar to that put forward for the case of aspartame, according to which all platyrrhine primates should be unable to detect these proteins as sweet due to a structural difference in the sweet-taste receptor, compared to catarrhine primates. Accordingly, previous studies on platyrrhine primate species report a general lack of ability of these species to detect these substances as sweet, whereas all catarrhine primates do so. Thus, the individual who showed a preference for monellin and thaumatin over water probably represents an exceptional case of variation in the sweet-taste receptor's typical alleles.
Sweet-tasting saccharides
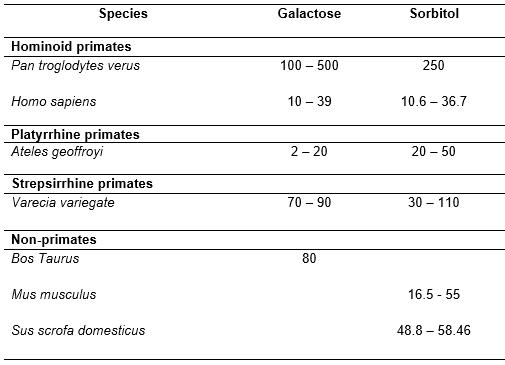
The spider monkeys' thresholds for galactose and sorbitol are lower than those of chimpanzees and similar to those of humans. This suggests a higher sensitivity of spider monkeys to this saccharide, as it is plausible to assume that spider monkeys may detect lower concentrations of a taste-substance than that indicated by their taste preference threshold.
As galactose is a constituent of lactose, a naturally occurring disaccharide in mammalian breastmilk, recent hypotheses have been put forward that associate the sensitivity to sweet-taste of a species to the experience of the sweet-taste of lactose in breastmilk. As for sorbitol, as this is a naturally-occurring saccharide in fruits, it is plausible to consider that the higher sensitivity of spider monkeys to this substance may be attributable to their higher degree of frugivory compared to other less sensitive species.
Spider monkeys’ perception of aspartame as indicated through facial responses
Taking together the results which are in line with those of previous studies, two distinct patterns of responsiveness to taste stimuli can be described, one being associated with presumably pleasant stimuli and the other being associated with presumably unpleasant stimuli. Thus, responses to pleasant stimuli include high frequencies of licking, sucking, and tongue protruding as well as slightly closed eyes. On the other hand, responses to aversive stimuli reveal lower frequencies of licking, sucking and tongue protruding, as well as higher frequencies of withdrawal from the taste stimuli.
Given the intermediate frequencies of these behaviors in response to aspartame, the results of the present study do not allow for an unequivocal determination of the spider monkeys’ perception of this substance. However, the results do suggest that aspartame is probably not perceived as sweet by spider monkeys. Rather, it is most likely perceived as either neutral or as slightly bitter, as indicated by similarities with, respectively, water and caffeine in the recorded frequencies for some of the analyzed behaviors.
Responsible for this page:
Director of undergraduate studies Biology
Last updated:
05/25/20